Innehåll
DNA är en av de få kombinationerna av bokstäver i kärnan i en vetenskaplig disciplin som verkar skapa en betydande nivå av förståelse även hos personer med liten livslängd exponering för biologi eller vetenskapen i allmänhet. De flesta vuxna som hör uttrycket "Its in her DNA" inser omedelbart att en viss egenskap är oskiljbar från den person som beskrivs; att kännetecknet på något sätt är född, aldrig försvinner och kan överföras till den personens barn och därefter. Detta verkar gälla även i sinnet hos dem som inte har någon aning om vad "DNA" ens står för, vilket är "deoxiribonukleinsyra."
Människor är förståeligt fascinerade av konceptet att ärva drag från sina föräldrar och överföra sina egna drag till sina avkommor. Det är bara naturligt för människor att fundera över sin egen biokemiska arv, även om få kan föreställa sig det så formellt. Erkännandet av att små osynliga faktorer i var och en av oss styr hur folk ser ut och till och med uppträder har säkert varit närvarande i många hundratals år. Men inte förrän i mitten av 1900-talet avslöjade modern vetenskap i härlig detalj inte bara hur de molekyler som var ansvariga för arv, utan också hur de såg ut.
Deoxyribonukleinsyra är verkligen den genetiska blått som alla levande saker upprätthåller i sina celler, en unik mikroskopisk finger som inte bara gör varje människa till en bokstavlig enskild individ (identiska tvillingar undantagna för nuvarande syften) utan avslöjar mycket viktigt information om varje person, från sannolikheten för att vara släkt med en annan specifik person till chansen att utveckla en viss sjukdom senare i livet eller överföra en sådan sjukdom till kommande generationer. DNA har blivit inte bara den naturliga centrala punkten för molekylärbiologi och livsvetenskap som helhet, utan också en integrerad del av kriminalteknik och biologisk teknik.
Upptäckten av DNA
James Watson och Francis Crick (och mindre vanligt, Rosalind Franklin och Maurice Wilkins) krediteras allmänt med upptäckten av DNA 1953. Denna uppfattning är dock felaktig. Kritiskt sett konstaterade dessa forskare faktiskt att DNA finns i tredimensionell form i form av en dubbel spiral, som i huvudsak är en stege vridna i olika riktningar i båda ändarna för att skapa en spiralform. Men dessa bestämda och ofta berömda forskare bygger "bara" på det noggranna arbetet för biologer som slet på jakt efter samma allmänna information redan 1860-talet, experiment som var lika banbrytande i sin egen rätt som Watsons, Crick och andra i forskningsperioden efter andra världskriget.
År 1869, 100 år innan människor skulle resa till månen, försökte en schweizisk kemist vid namn Friedrich Miescher att utvinna proteinkomponenterna från leukocyter (vita blodkroppar) för att bestämma deras sammansättning och funktion. Det han istället extraherade kallade han "nuklein", och även om han saknade de instrument som behövdes för att lära sig vad framtida biokemister skulle kunna lära sig, upptäckte han snabbt att denna "nuklein" var relaterad till proteiner men inte själv var protein, att den innehöll en ovanlig mängd fosfor, och att det här ämnet var resistent mot att brytas ned av samma kemiska och fysiska faktorer som nedbrytade proteiner.
Det skulle gå över 50 år innan den verkliga betydelsen av Mieschers arbete först blev uppenbar. Under det andra decenniet av 1900-talet var en rysk biokemist, Phoebus Levene, den första som föreslog att, det vi kallar nukleotider idag, bestod av en sockerdel, en fosfatdel och en basdel; att sockret var ribos; och att skillnaderna mellan nukleotider var skyldiga skillnaderna mellan deras baser. Hans "polynukleotid" -modell hade några brister, men enligt dagens normer var den anmärkningsvärt i mål.
1944 var Oswald Avery och hans kollegor vid Rockefeller University de första kända forskarna som formellt antydde att DNA bestod av ärftliga enheter eller gener. Efter att ha följt upp såväl som hos Levene gjorde den österrikiska forskaren Erwin Chargaff två viktiga upptäckter: en, att sekvensen av nukleotider i DNA varierar mellan organismer, i motsats till vad Levene hade föreslagit; och två att den totala mängden kvävebaser adenin (A) och guanin (G) kombinerad, oavsett art, var i stort sett alltid samma som den totala mängden cytosin (C) och timin (T). Detta ledde inte riktigt till att Chargaff drog slutsatsen att A-par med T- och C-par med G i allt DNA, men det hjälpte senare att stärka den slutsats som andra når.
Slutligen, 1953, Watson och hans kollegor, som gynnades av snabbt förbättrade sätt att visualisera tredimensionella kemiska strukturer, sammansatte alla dessa fynd och använde kartongmodeller för att fastställa att en dubbel spiral passade allt som var känt om DNA på något sätt ingenting annars kunde.
DNA och ärftliga drag
DNA identifierades som det ärftliga materialet i livings-saker långt innan dess struktur klargjordes, och som ofta fallet i experimentvetenskapen var denna vitala upptäckt faktiskt en anledning till forskarnas huvudsyfte.
Innan antibiotikabehandling uppstod i slutet av 1930-talet, krävde infektionssjukdomar mycket mer människoliv än de gör idag, och att avslöja mysterierna för de ansvariga organismerna var ett kritiskt mål i mikrobiologisk forskning. 1913 påbörjade ovannämnda Oswald Avery arbete som i slutändan avslöjade ett högt polysackaridinnehåll (socker) i kapslar av pneumokockbakterier som hade isolerats från lunginflammationspatienter. Avery teoretiserade att dessa stimulerade produktionen av antikroppar hos infekterade människor. Under tiden i England utförde William Griffiths arbete som visade att döda komponenter av en typ av sjukdom-orsakande pneumokocker kunde blandas med de levande komponenterna i en ofarlig pneumokock och producera en sjukdom-orsakande form av den tidigare ofarliga typen; detta bevisade att allt som rörde sig från de döda till de levande bakterierna var ärftligt.
När Avery fick reda på Griffiths resultat, började han genomföra reningsexperiment i ett försök att isolera det exakta materialet i de pneumokocker som var ärftliga och hängde in på nukleinsyror, eller mer specifikt, nukleotider. DNA var redan starkt misstänkt för att ha vad som då populärt kallades "omvandlingsprinciper", så Avery och andra testade denna hypotes genom att utsätta det ärftliga materialet för en mängd olika medel. De som var kända för att vara destruktiva för DNA-integritet men ofarliga för proteiner eller DNA, kallade DNAaser, var tillräckliga i stora mängder för att förhindra överföring av egenskaper från en bakteriegeneration till nästa. Under tiden skadade proteaser, som avlägsnar proteiner.
Hemmet till Averys och Griffiths arbete är att återigen, medan människor som Watson och Crick med rätta har lovordats för sina bidrag till molekylär genetik, var etablering av DNA-strukturen faktiskt ett ganska sent bidrag till processen att lära sig om detta spektakulär molekyl.
Strukturen för DNA
Chargaff, även om han uppenbarligen inte beskrev strukturen för DNA i sin helhet, visade att förutom (A + G) = (C + T), de två strängarna som kändes för att ingå i DNA alltid var samma avstånd från varandra. Detta ledde till postulatet som puriner (inklusive A och G) alltid bundna till pyrimidiner (inklusive C och T) i DNA. Detta gjorde tredimensionell mening, eftersom puriner är betydligt större än pyrimidiner, medan alla puriner är väsentligen samma storlek och alla pyrimidiner är väsentligen samma storlek. Detta antyder att två puriner bundna tillsammans skulle ta upp betydligt mer utrymme mellan DNA-strängar än två pyrimidiner, och att varje given purin-pyrimidinparring skulle konsumera samma mängd utrymme. Att lägga all denna information krävde att A binder till och endast till T och att samma förhållande gäller för C och G om denna modell skulle visa sig framgångsrik. Och det har det.
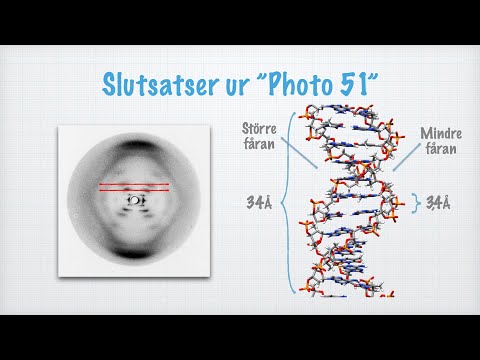
Baserna (mer om dessa senare) binder till varandra på insidan av DNA-molekylen, som rullar i en stege. Men hur är det med strängarna eller "sidorna" själva? Rosalind Franklin, som arbetade med Watson och Crick, antog att denna "ryggrad" var gjord av socker (specifikt ett pentosesocker, eller en med en 5-atoms ringstruktur) och en fosfatgrupp som länkar socker. På grund av den nyligen klarade idén om basparning, blev Franklin och de andra medvetna om att de två DNA-strängarna i en enda molekyl var "komplementära", eller i själva verket spegelbilder av varandra på nivån av deras nukleotider. Detta gjorde det möjligt för dem att förutsäga den ungefärliga radien för den tvinnade formen av DNA inom en solid grad av noggrannhet, och röntgendiffraktionsanalys bekräftade den spiralformade strukturen. Idén att helixen var en dubbel spiral var den sista stora detaljen om DNA: s struktur som föll på plats 1953.
Nukleotider och kvävebaser
Nukleotider är de upprepande underenheterna av DNA, vilket är motsatsen till att säga att DNA är en polymer av nukleotider. Varje nukleotid består av ett socker som kallas deoxiribos som innehåller en femkantig ringstruktur med en syre och fyra kolmolekyler. Detta socker är bundet till en fosfatgrupp, och två fläckar längs ringen från denna position är också bundna till en kvävehaltig bas. Fosfatgrupperna sammanlänker sockret för att bilda DNA-ryggraden, vars två strängar vrider sig runt de bundna kvävtunga baserna i mitten av den dubbla spiralen. Helixen gör en komplett 360-graders twist ungefär var 10 baspar.
Ett socker som endast är bundet till en kvävehaltig bas kallas a nukleosid.
RNA (ribonukleinsyra) skiljer sig från DNA på tre viktiga sätt: En, pyrimidin uracil ersätter tymin. Två, pentosesocker är ribos snarare än deoxiribos. Och tre är RNA nästan alltid enkelsträngad och finns i flera former, diskussionen som ligger utanför denna artikel.
DNA-replikation
DNA "packas upp" i sina två komplementära strängar när det är dags att kopior ska göras. När detta händer bildas dottersträngar längs ensamstående föräldrasträngar. En sådan dottersträng bildas kontinuerligt genom tillsats av enstaka nukleotider, under påverkan av enzymet DNA-polymeras. Denna syntes följer helt enkelt längs riktningen för separationen av moder-DNA-strängarna. Den andra dottersträngen bildas från små polynukleotider som kallas Okazaki-fragment som faktiskt bildas i motsatt riktning för att packa upp föräldrasträngar, och sedan sammanfogas av enzymet DNA-ligas.
Eftersom de två dottersträngarna också är komplementära till varandra, binds deras baser så småningom för att göra en dubbelsträngad DNA-molekyl identisk med den överordnade.
I bakterier, som är encelliga och kallas prokaryoter, sitter en enda kopia av bakterias DNA (även kallad genomet) i cytoplasma; ingen kärna finns. I multicellulära eukaryota organismer finns DNA i kärnan i form av kromosomer, som är starkt spiralformade, spolade och rumsligt kondenserade DNA-molekyler som bara är miljondelar av en meter lång och proteiner kallas histoner. Vid mikroskopisk undersökning liknar de kromosomdelarna som visar växlande histon "spolar" och enkla DNA-strängar (kallad kromatin på denna organisationsnivå) till pärlor på en sträng. En del eukaryot DNA finns också i organeller av celler som kallas mitokondrier.